
Mechanism of Cryptotanshinone Inhibiting Proliferation of Human Breast Cancer MCF7 Cells
-
摘要:目的
探讨隐丹参酮(CPT)对人乳腺癌MCF7细胞的抑制作用及机制。
方法MTT法检测CPT干预后MCF7细胞存活率,Annexin V/PI实验、Hoechst33258荧光染色实验检测细胞凋亡;流式细胞术检测细胞周期及细胞内活性氧ROS;细胞划痕实验、Transwell小室实验检测细胞迁移与侵袭;微球体培养及流式细胞术检测微球体表面分子CD44与CD24;Western blot检测细胞相关蛋白的表达。
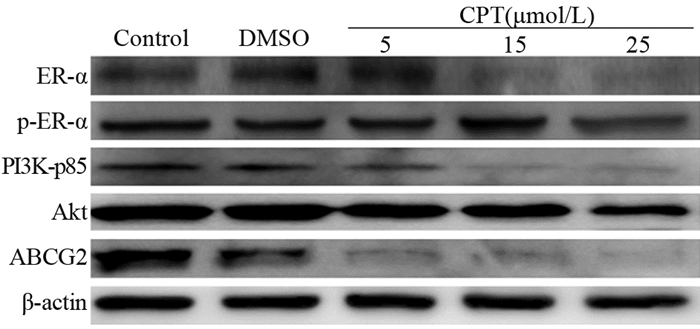
结果CPT呈剂量依赖性地抑制MCF7细胞增殖,24 h半数抑制浓度(IC50)为19.24 μmol/L。与未加药处理组相比,CPT处理组可将细胞周期阻滞于S期,并诱导细胞凋亡。细胞划痕实验和Transwell小室结果表明,CPT对MCF7细胞的迁移与侵袭有显著抑制作用。此外,CPT降低了MCF7细胞来源微球体中CD24−/lowCD44+细胞群。Western blot结果证明,CPT能明显上调Bax蛋白表达,下调Bcl2、PI3K-p85、Akt、N-cadherin、Twist1、Sox2、Oct4、Nanog蛋白表达,有效抑制ER-α磷酸化,降低了多重药物转运蛋白ABCG2的表达。
结论CPT通过抑制MCF7细胞迁移与侵袭、降低CD24−/low CD44+细胞群数量,影响肿瘤干细胞相关蛋白表达降低细胞干性,从而抑制细胞增殖。
Abstract:ObjectiveTo investigate the inhibitory effect of cryptotanshinone (CPT) on human breast cancer cell MCF7 and its mechanism.
MethodsThe survival rate of MCF7 cells was measured by MTT assay. Cell apoptosis was detected by Annexin V/PI assay and Hoechst 33258 fluorescence staining assay. Cell cycle and reactive oxygen species were detected by flow cytometry. Cell migration and invasion were detected by cell scratch test and Transwell chamber test. The surface molecules CD44 and CD24 were detected by flow cytometry and microsphere culture. The expression of cell-associated proteins was detected by Western blot.
ResultsCPT inhibited the proliferation of MCF7 cells in a dose-dependent manner, and the 24 h IC50 value was 19.24 μmol/L. Compared with the untreated group, the CPT-treated group showed cell cycle arrested in the S phase, and apoptosis was induced. The results of the cell scratch and Transwell chamber tests showed that CPT significantly inhibited the migration and invasion of MCF7 cells. Furthermore, CPT reduced the CD24-/LowCD44+ cell population in MCF7 cell-derived microspheres. Western blot results showed that CPT could up-regulate the expression of Bax protein, down-regulate the expression of BCL-2, PI3K-p85, Akt, N-cadherin, Twist1, Sox2, Oct4, and Nanog protein, effectively inhibit the phosphorylation of ER-α, and decrease the expression of ABCG2.
ConclusionCPT can inhibit the proliferation of MCF7 cells by inhibiting the migration and invasion of MCF7 cells, decreasing the number of CD24-/lowCD44+ cells and affecting the expression of tumor stem cell-related proteins.
-
Key words:
- Cryptotanshinone /
- Breast cancer /
- Migration and invasion /
- Tumor stem cell /
- Mechanism of action
-
0 引言
结肠癌是全球最常见的恶性肿瘤之一,手术切除是目前治疗结肠癌的首选方法,但仍有50%~60%的根治性术后患者会出现复发或远处转移[1],预后差,复发率高是结肠癌恶性度高的主要原因。因此寻找结肠癌远期预后的特异性指标显得尤为重要。近年来,错配修复基因(mismatch repair gene, MMR)成为肿瘤研究热点,由于DNA错配修复基因种系突变可导致区域内体细胞突变高累积,故又称为高度微卫星不稳定(MSI-H),目前公认两者等同(下文统称deficient mismatch repair, dMMR)[2]。dMMR在Ⅱ/Ⅲ期结肠癌中被证明具有明显的预后优势,是结肠癌治疗中重要的生物标志物及预后因子[3]。另外中性粒细胞/淋巴细胞比率(neutrophil lymphocyte ratio, NLR)作为全身炎性反应状态的有用指标,被认为可反映促肿瘤炎性反应和抗肿瘤免疫功能之间的平衡,其预后意义已在结肠癌等多种实体瘤中得到广泛证实[4]。研究表明dMMR结肠癌表现出更高水平的肿瘤浸润淋巴细胞(tumor infiltrating lymphocyte, TIL)及良好的肿瘤微环境,这可能是其预后良好的主要机制[5]。但是鲜有研究在MMR的背景下研究机体全身炎性反应状态,dMMR状态下的局部炎性反应浸润与全身的炎性反应状态的联系尚无确切的定论。因此通过挖掘MMR与NLR的相关性,或许可以进一步阐述dMMR结肠癌的预后机制。
1 资料与方法
1.1 临床资料
回顾性收集2015年4月1日—2017年4月1日期间在福建医科大学附属第二医院和厦门大学附属第一医院接受左/右半结肠癌根治术、术后病理确诊为结肠癌的125名患者资料,其中优先纳入免疫组织化学证实为dMMR状态结肠癌55例,并按标准随机纳入70例错配修复基因正常(proficient mismatch repair, pMMR)状态结肠癌用作对比。纳入标准:(1)结肠癌得到病理证实,术后病理进行错配修复蛋白免疫组织化学分析;(2)术前均完善了血常规、生化、肿瘤标志物检查;(3)术前未接受新辅助放化疗及其他辅助治疗。排除标准:(1)伴有第2种原发性恶性肿瘤;(2)术前明确合并急性、慢性感染或自身免疫性疾病等;(3)随访期间因其他疾病或事件死亡。临床资料包括年龄、性别、肿瘤病理特征(大小、部位、大体类型、组织学类型、分化程度、浸润深度、淋巴结转移、分期)、CEA、乳酸脱氢酶、术前NLR、术后接受化疗情况。本研究均经所有患者知情同意并获得授权签字(伦理号:[2019]福医附二伦理审字第(116号))。
1.2 免疫组织化学结果判定
免疫组织化学法检测肿瘤组织中错配修复基因表达情况。术后病理报告经高年资病理科医师审核确认。错配修复蛋白MLH1、PMS2、MSH2、MSH6中出现1个及以上表达缺失即判定为dMMR状态,未出现错配修复蛋白缺失即判定为pMMR状态。
1.3 NLR值
结肠癌根治术前3天内所有患者静脉采血进行血常规检查。NLR为中性粒细胞绝对值和淋巴细胞绝对值的比值。确保所有患者术前无明显的感染症状。
1.4 观察指标
复发标准为:本院或外院影像学检查(B型超声、CT、MRI、胸片、肠镜等)发现病灶或者直接确诊的病理证据。主要研究指标是无复发生存期(recurrence-free survival, RFS)。
1.5 随访情况
所有患者均通过门诊、电话、微信方式进行随访,随访终点为患者结肠癌复发。数据收集截至2020年4月1日,过程中失访及未复发患者作为结尾数据处理。
1.6 统计学方法
采用SPSS23.0软件作统计学数据分析。ROC曲线确定NLR最佳阈值。计数资料采用卡方检验。Kaplan-Meier法计算不同MMR状态及NLR组别术后无病生存率和累计生存率,Log rank检验比较各组别生存时间的差异。P < 0.05为差异有统计学意义。
2 结果
2.1 结肠癌患者的一般资料
共125例患者纳入研究,其中男性65例,女性60例。中位年龄63岁(33~90岁)。按患者MMR状态分为dMMR组(55例),pMMR组(70例)。
2.2 不同MMR状态组临床病理特征分析
相比pMMR组,dMMR组结肠癌发病年龄较低,肿瘤体积更大,分化程度更低,多发于右半结肠,更多见神经侵犯(均P < 0.05)。在性别、大体类型、组织类型、浸润深度、淋巴结转移、病理分期、CEA、乳酸脱氢酶上两组间差异无统计学意义(均P > 0.05),见表 1。
表 1 结肠癌患者临床资料(n(%))Table 1 Clinical data of colon cancer patients (n(%))
2.3 MMR状态与术前NLR相关性及预后价值分析
根据术前NLR值和患者肿瘤复发情况建立ROC曲线,患者NLR值ROC曲线下面积为0.644。根据兼顾敏感度和特异性的原则,最终确定NLR=3作为截断值,见图 1。将NLR < 3定义为低NLR值,NLR≥3定义为高NLR值。最终结果显示,dMMR组(n=55)中低NLR值40例,高NLR值15例(72.7% vs. 27.3%);pMMR组(n=70)中低NLR值30例,高NLR值40例(42.9% vs. 57.1%)。两组差异有统计学意义(P < 0.05)。
125例患者随访36月。术后根据NCCN指南,共77例患者接受术后辅助化疗,dMMR组36例,pMMR组41例(65.5% vs. 58.6%)(P > 0.05)。随访过程中9例患者未能获得完整的随访资料,将其纳入结尾数据分析。结果3年总复发率31.2%。按MMR状态和NLR高低,将患者分为四组(分别为dMMR并低NLR组,dMMR并高NLR组,pMMR并低NLR组,pMMR并高NLR组),四组患者数量,3年无复发生存率比较,见表 2。组间进行卡方检验,结果显示,四组中,dMMR并低NLR组3年无复发生存率明显较高(P < 0.05)。Kaplan-Meier生存分析,结果见图 2,组间采用Log rank检验,结果见表 3。对比pMMR并高NLR组,dMMR并低NLR组存在明显的生存优势(P < 0.05);而dMMR并高NLR组、pMMR并低NLR组两组接近(P > 0.05)。
表 2 不同MMR状态和NLR水平组的3年无复发生存率情况比较(n(%))Table 2 Comparison of 3-year relapse-free survival rates among groups with different MMR status and NLR level (n(%))
![]() 图 2 不同MMR状态和NLR水平组的生存分析曲线Figure 2 Survival curves of different MMR status and NLR level groups表 3 不同MMR状态和NLR水平组的Log rank检验结果Table 3 Log rank statistics result of different MMR status and NLR level groups
图 2 不同MMR状态和NLR水平组的生存分析曲线Figure 2 Survival curves of different MMR status and NLR level groups表 3 不同MMR状态和NLR水平组的Log rank检验结果Table 3 Log rank statistics result of different MMR status and NLR level groups
3 讨论
DNA错配修复基因系统主要包括4个蛋白(MLH1,PMS2,MSH2,MSH6), 出现1个及以上表达缺失即判定为dMMR。在亚洲人群中,dMMR结肠癌占全部结直肠癌的5.5%~7.3%[2, 6]。近年来,许多的研究皆证实了dMMR结肠癌独特的预后优势,一项Meta分析评估了1 277例Ⅰ~Ⅳ期dMMR结肠癌患者,发现无论早期还是进展期疾病,dMMR复发率明显低于pMMR肿瘤,减少死亡风险35%[7]。在对其机制的探索中,研究发现了dMMR结肠癌独特的病理学特征。例如对比pMMR结肠癌,dMMR患者年龄偏低,肿瘤好发于右半结肠,癌灶病理分化程度差,多见黏液分泌[8]。这本应决定较差的预后结果,逆转的关键在于dMMR结肠癌在宿主-肿瘤相互作用上有更明显的肿瘤浸润特征,如淋巴反应,克罗恩样淋巴结节,包括更多的生发中心和上皮内淋巴细胞浸润[9],这决定了良好的肿瘤浸润微环境,患者表现更长的总生存期(overall survival, OS)、无复发生存期(recurrence-free survival, RFS)[10]。中性粒细胞/淋巴细胞比值(NLR)作为一种反映机体炎性反应程度和免疫状态的有用指标,被发现与结直肠癌等肿瘤的复发和预后关系密切[3]。但据我们所知,很少有研究同时考虑肿瘤的局部和全身炎性反应,进一步评估其预后价值。有一项胃癌研究表明肿瘤基质中高密度的CD4+T细胞与低NLR相关,但遗憾的是该研究并没有表明MMR状态[11]。
本研究结果显示,MMR状态与患病年龄、肿瘤发生部位、肿瘤大小、肿瘤分化程度、癌灶神经侵犯相关。相比pMMR组,dMMR组患者患病年龄低,肿瘤多发生于右半结肠,且体积大,肿瘤分化程度低,伴随神经侵犯,这与国内赵喜连等[12]研究结果相同,区别在于本研究未能显示淋巴结转移、肿瘤浸润深度方面的相关性。但不管如何,两者均体现出dMMR结肠癌具有区别于pMMR的临床病理特征,表明对具有该病理特征的结肠癌更应进行MMR检测,以便提高dMMR筛查的敏感度和特异性,对指导患者的治疗具有重要意义。在对不同状态的术前NLR值分析后发现,dMMR与术前NLR明显相关,dMMR组低NLR的患者(72.7%)比例明显高于pMMR组(42.9%)。既往研究发现中性粒细胞衍生的细胞因子和趋化因子有利于肿瘤增殖,局部浸润和血管形成,中性粒细胞增加可加速局部肿瘤的侵袭和远处转移的形成,增加的肿瘤内淋巴细胞浓度可放大至宿主全身性炎性反应,可能与积极的临床结果相关,低NLR明显更有利于肿瘤预后[4, 13]。在本研究中,对MMR和NLR状态分组后进行复发率的随访,我们发现dMMR并低NLR组3年无复发生存率明显高于其他三组,生存分析显示相比pMMR高NLR组具有明显的术后生存优势。这提示dMMR低NLR是结肠癌的良好预后因素。关于dMMR状态和NLR的联系,我们猜测有以下两点:一是低NLR可能是良好肿瘤浸润微环境的全身性体现,两者互为因果,共同提示结肠癌的预后;二是可能存在更复杂的机制同时调控两者状态。这需要未来更多的研究来证实。
MMR状态和NLR对结肠癌的发生、发展具有重要意义,但数据中仍存在许多不足之处需加以解决。首先,目前关于NLR的界值尚没有明确定论,代表全身炎性反应状态的程度没有确切说法,既往研究也都是经验性的指标。根据最近的评论[14],该临界值介于2到5之间,其在不同的研究、不同的种族都可能有所不同,因此关于NLR的临床应用,这可能是一个关键限制。另外,本研究为回顾性研究,术前缺少NLR的基线值,研究群体样本量不足,多数为适合接受手术的非进展期患者,缺乏代表性,且术后随访复查视患者依从性而定,部分患者真实复发时间可能延后于随访所得时间,这些均是造成研究结果偏倚的重要因素;其次,关于全身炎性反应状态的指标有很多,包括C-反应蛋白(CRP)、IL家族细胞因子、预后营养指数(PNI),均有相关的文献支持其在肿瘤中的应用[15],但因为并不作为常规检测,因此并没有被纳入本研究,期待未来开展大型的前瞻性研究进一步弥补这些短板;最后,重要的一点是我们应认识到随着放化疗、靶向治疗、免疫治疗等多种新兴治疗手段的出现,结肠癌的预后模型构建也变得更加的复杂和困难,未来还有很长的路要走,需要我们共同努力。
综上,dMMR结肠癌具有独特的临床病理特征,联合NLR值可用于评估患者术后复发风险。根据其影响,应注意督促患者复查随访,及时采取干预手段,降低结肠癌死亡率,提高患者生存质量。
Competing interests: The authors declare that they have no competing interests.利益冲突声明:所有作者均声明不存在利益冲突。作者贡献:杨舒涵:研究设计、文章撰写及修改王玉琴:初稿撰写刘航:实验实施夏丽洁:初稿修改刘苏颖:协助整理数据,补充研究设计缺陷张英:文章审阅及修改 -
![]()
图 1 MCF7细胞经CPT干预后细胞形态(A, ×200)与细胞活力变化(B)
Figure 1 Cell morphology(A, ×200) and cell viability(B) of MCF7 cells after CPT intervention
![]()
图 2 CPT干预下MCF7细胞凋亡现象的观察:流式细胞术(A)、Western blot(B)及hoechst 33258染色(C, ×200)
Figure 2 Apoptosis of MCF7 cells under CPT intervention observed by flow cytometry(A), Western blot(B) and hoechst 33258 staining(C, ×200)
![]()
图 3 流式细胞检测CPT可引起MCF7细胞周期阻滞
Figure 3 CPT induced cell cycle arrest in MCF7 cells detected by flow cytometry
![]()
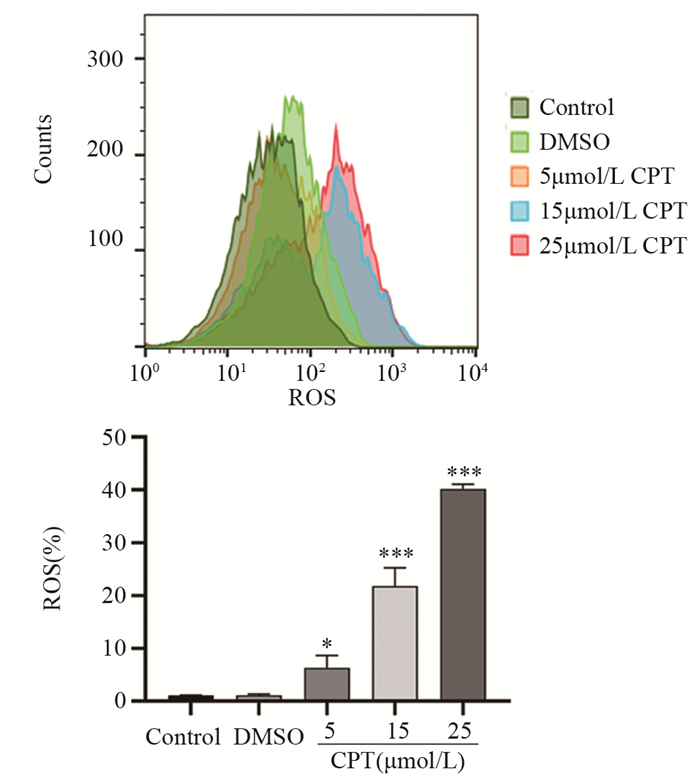
图 4 流式细胞检测显示CPT诱导MCF7细胞ROS产生
Figure 4 CPT induced generation of intracellular ROS in MCF7 cells detected by flow cytometry
![]()
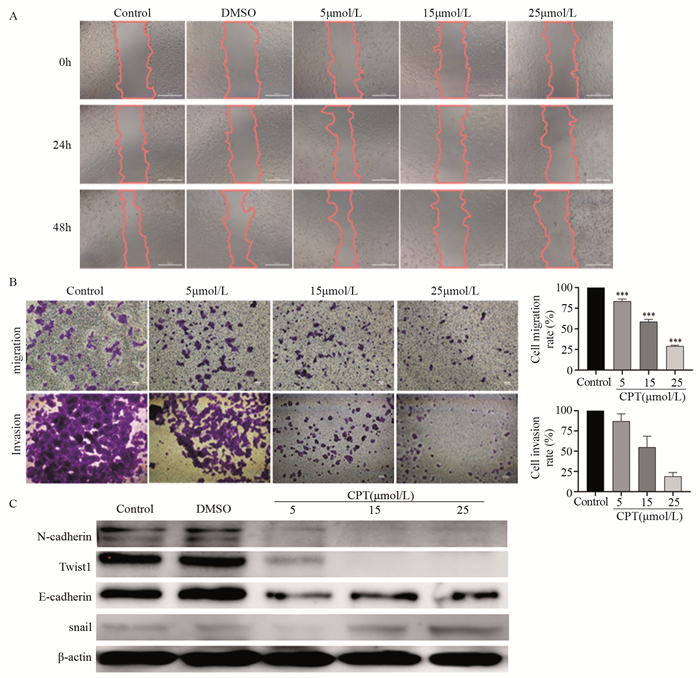
图 5 CPT对MCF7细胞迁移与侵袭能力的影响
Figure 5 Effect of CPT on migration and invasion abilities of MCF7 cells detected
![]()
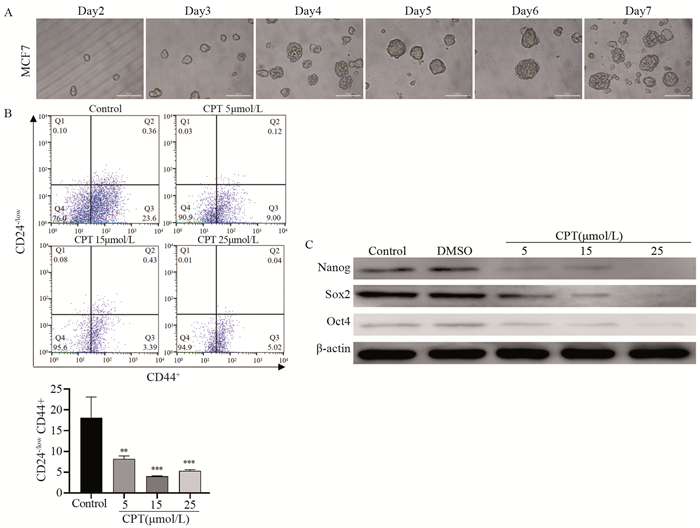
图 6 MCF7细胞来源微球体培养(A)、微球体CD24-/low CD44+表达情况(B)及肿瘤干细胞相关蛋白表达(C)
Figure 6 Culture of tumor spheres derived from MCF7 cells(A) and tumor spheres detection of CD24-/lowCD44+expression(B) and expression of cancer stem cell-related proteins(C)
-
[1] Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209-249. doi: 10.3322/caac.21660
[2] Schmid P, Salgado R, Park YH, et al. Pembrolizumab plus chemotherapy as neoadjuvant treatment of high-risk, early-stage triple-negative breast cancer: results from the phase 1b open-label, multicohort KEYNOTE-173 study[J]. Ann Oncol, 2020, 31(5): 569-581. doi: 10.1016/j.annonc.2020.01.072
[3] Hanker AB, Sudhan DR, Arteaga CL. Overcoming Endocrine Resistance in Breast Cancer[J]. Cancer Cell, 2020, 37(4): 496-513. doi: 10.1016/j.ccell.2020.03.009
[4] Tang JN, Wu ZY, Tian ZL, et al. OTUD7B stabilizes estrogen receptor α and promotes breast cancer cell proliferation[J]. Cell Death Dis, 2021, 12(6): 534. doi: 10.1038/s41419-021-03785-7
[5] Kim H, Park SH, Lee J, et al. TNFα Enhances Tamoxifen Sensitivity through Dissociation of ERα-p53-NCOR1 Complexes in ERα- Positive Breast Cancer[J]. Cancers(Basel), 2021, 13(11): 2601.
[6] Kong D, Zhou H, Neelakantan D, et al. VEGF-C mediates tumor growth and metastasis through promoting EMT-epithelial breast cancer cell crosstalk[J]. Oncogene, 2021, 40(5): 964-979. doi: 10.1038/s41388-020-01539-x
[7] Bae MJ, Karadeniz F, Oh JH, et al. MMP-Inhibitory Effects of Flavonoid Glycosides from Edible Medicinal Halophyte Limonium tetragonum[J]. Evid Based Complementary Alternat Med, 2017, 2017: 6750274.
[8] Sun B, Lin Y, Wang X, et al. Single Nucleotide Polymorphism of the Enhancer of Zeste Homolog 2 Gene rs2072408 is Associated with Lymph Node Metastasis and Depth of Primary Tumor Invasion in Gastric Cancer[J]. Clini Lab, 2016, 62(11): 2099-2105.
[9] Elkashty OA, Abu Elghanam G, Su X, et al. Cancer stem cells enrichment with surface markers CD271 and CD44 in human head and neck squamous cell carcinomas[J]. Carcinogenesis, 2020, 41(4): 458-466. doi: 10.1093/carcin/bgz182
[10] Koch C, Kuske A, Joosse SA, et al. Characterization of circulating breast cancer cells with tumorigenic and metastatic capacity[J]. EMBO Mol Med, 2020, 12(9): e11908. doi: 10.15252/emmm.201911908
[11] Zeng X, Liu C, Yao J, et al. Breast cancer stem cells, heterogeneity, targeting therapies and therapeutic implications[J]. Pharmacol Res, 2021, 163: 105320. doi: 10.1016/j.phrs.2020.105320
[12] Wang KH, Kao AP, Lin TC, et al. Promotion of epithelial– mesenchymal transition and tumor growth by 17β-estradiol in an ER+/HER2+ cell line derived from human breast epithelial stem cells[J]. Biotechnol Appl Biochem, 2012, 59(3): 262-267. doi: 10.1002/bab.1022
[13] Semina SE, Alejo LH, Chopra S, et al. Identification of a novel ER-NFĸB-driven stem-like cell population associated with relapse of ER+ breast tumors[J]. Breast Cancer Res, 2022, 24(1): 88. doi: 10.1186/s13058-022-01585-1
[14] Yan Y, Huang H. Interplay Among PI3K/AKT, PTEN/FOXO and AR Signaling in Prostate Cancer[J]. Adv Exp Med Biol, 2019, 1210: 319-331.
[15] Zattoni IF, Delabio LC, Dutra JP, et al. Targeting breast cancer resistance protein (BCRP/ABCG2): Functional inhibitors and expression modulators[J]. Eur J Medl Chem, 2022, 237: 114346. doi: 10.1016/j.ejmech.2022.114346
[16] 李铮, 侯炜. 隐丹参酮对肺鳞癌KAL△LU细胞增殖和周期以及干细胞抗原1的影响[J]. 环球中医药, 2022, 15(4): 577-582. https://www.cnki.com.cn/Article/CJFDTOTAL-HQZY202204005.htm Li Z, Hou W. The effects of Cryptotanshinone on cell proliferation and cell cycle and Sca-1 of lung squamous KALΔLU cells[J]. Huan Qiu Zhong Yi Yao, 2022, 15(4): 577-582. https://www.cnki.com.cn/Article/CJFDTOTAL-HQZY202204005.htm
[17] Li H, Gao C, Liu C, et al. A review of the biological activity and pharmacology of cryptotanshinone, an important active constituent in Danshen[J]. Biomed Pharmacother, 2021, 137: 111332. doi: 10.1016/j.biopha.2021.111332
[18] Zhang Q, Wang L, Gan C, et al. Cryptotanshinone Induces Apoptosis and Inhibits Migration and Invasion in Human Hepatocellular Carcinoma Cells In Vitro[J]. Nat Prod Commun, 2020, 15(1): 1934578X1989957.
[19] Park M, Kim D, Ko S, et al. Breast Cancer Metastasis: Mechanisms and Therapeutic Implications[J]. Int J Mol Sci, 2022, 23(12): 6806. doi: 10.3390/ijms23126806
[20] Yen JH, Huang HS, Chuang CJ, et al. Activation of dynaminrelated protein 1-dependent mitochondria fragmentation and suppression of osteosarcoma by cryptotanshinone[J]. J Exp Clin Cancer Res, 2019, 38(1): 42. doi: 10.1186/s13046-018-1008-8
[21] Francou A, Anderson KV. The Epithelial-to-Mesenchymal Transition(EMT) in Development and Cancer[J]. Annu Rev Cancer Biol, 2020, 4: 197-220. doi: 10.1146/annurev-cancerbio-030518-055425
[22] Sameri S, Saidijam M, Bahreini F, et al. Cancer Chemopreventive Activities of Silibinin on Colorectal Cancer through Regulation of E-Cadherin/β-Catenin Pathway[J]. Nutr Cancer, 2021, 73(8): 1389-1399. doi: 10.1080/01635581.2020.1800764
[23] Padmanaban V, Krol I, Suhail Y, et al. E-cadherin is required for metastasis in multiple models of breast cancer[J]. Nature, 2019, 573(7774): 439-444. doi: 10.1038/s41586-019-1526-3
[24] Elisha Y, Kalchenko V, Kuznetsov Y, et al. Dual role of E-cadherin in the regulation of invasive collective migration of mammary carcinoma cells[J]. Sci Rep, 2018, 8(1): 4986. doi: 10.1038/s41598-018-22940-3
[25] Cai F, Xiao H, Sun Y, et al. Expression of Snail and E-cadherin in Drug-resistant MCF-7/ADM Breast Cancer Cell Strains[J]. J Coll Physicians Surg Pak, 2019, 29(3): 240-244. doi: 10.29271/jcpsp.2019.03.240
[26] Saénz-de-Santa-María I, Celada L, Chiara MD. The Leader Position of Mesenchymal Cells Expressing N-Cadherin in the Collective Migration of Epithelial Cancer[J]. Cells, 2020, 9(3): 731. doi: 10.3390/cells9030731
[27] Najafi M, Farhood B, Mortezaee K. Cancer stem cells (CSCs) in cancer progression and therapy[J]. J Cell Physiol, 2019, 234(6): 8381-8395. doi: 10.1002/jcp.27740
[28] Ervin EH, French R, Chang CH, et al. Inside the stemness engine: Mechanistic links between deregulated transcription factors and stemness in cancer[J]. Semin Cancer Biol, 2022, 87: 48-83. doi: 10.1016/j.semcancer.2022.11.001
[29] Zhou C, Wang D, Li J, et al. TGFB2-AS1 inhibits triple-negative breast cancer progression via interaction with SMARCA4 and regulating its targets TGFB2 and SOX2[J]. Proc Natl Acad Sci U S A, 2022, 119(39): e2117988119. doi: 10.1073/pnas.2117988119
[30] Xu LM, Zhang J, Ma Y, et al. MicroRNA-135 inhibits initiation of epithelial-mesenchymal transition in breast cancer by targeting ZNF217 and promoting m6A modification of NANOG[J]. Oncogene, 2022, 41(12): 1742-1751. doi: 10.1038/s41388-022-02211-2
[31] Pancholi S, Leal MF, Ribas R, et al. Combination of mTORC1/2 inhibitor vistusertib plus fulvestrant in vitro and in vivo targets oestrogen receptor-positive endocrine-resistant breast cancer[J]. Breast Cancer Res, 2020, 21(1): 14.
[32] Huang CY, Deng JS, Huang WC, et al. Attenuation of Lipopolysaccharide-Induced Acute Lung Injury by Hispolon in Mice, Through Regulating the TLR4/PI3K/Akt/mTOR and Keap1/Nrf2/HO-1 Pathways, and Suppressing Oxidative Stress-Mediated ER Stress-Induced Apoptosis and Autophagy[J]. Nutrients, 2020, 12(6): 1742. doi: 10.3390/nu12061742
[33] Xie ZY, Liu MS, Zhang C, et al. Aspirin enhances the sensitivity of hepatocellular carcinoma side population cells to doxorubicin via miR-491/ABCG2[J]. Biosci Rep, 2018, 38(6): BSR20180854. doi: 10.1042/BSR20180854
[34] Dakhlaoui I, Vahdati S, Maalej E, et al. Synthesis and biological assessment of new pyrimidopyrimidines as inhibitors of breast cancer resistance protein (ABCG2)[J]. Bioorg Chem, 2021, 116: 105325. doi: 10.1016/j.bioorg.2021.105325
-
期刊类型引用(0)
其他类型引用(1)
 下载:
下载:





计量
- 文章访问数: 2080
- HTML全文浏览量: 2954
- PDF下载量: 529
- 被引次数: 1

